 |
Evaluation of interspecific diversity in Avena genus by RAPD analysis |
| Dr. Igor Loskutov |
 |
Till our days systematic position of the species in genus Avena L. still remains disputable among the researchers. Two tendencies are obvious in the modern classifications of Avena L.: expansion of the volume of species (so-called biological species) and subdivision of species into smaller ones on the basis of morphological and especially karyological differences. Extreme examples of these trends are such classifications where the number of species is either only 14, or amounts to 34. In this connection a molecular approach was taken to the study of phylogenetic relationships of Avena diploid and polyploid species based on RAPD-markers analyses (Randomy Amplified Polymorphic DNA).
Materials and Methods.
In this investigation (Perchuk et al,. 1999, joint project VIR (Russia) – NIAR (Japan) genetic variability in RAPDs was studied in 74 accessions mostly Mediterranean origin represented 20 species with different ploidy level – di-, tetra- and hexaploids. They are: diploids – A.clauda Dur., A.pilosa M.B. (Cp genome); A.ventricosa Bal. (Cv genome); A.longiglumis Dur. (Al genome); A.canariensis Baum (Ac genome); A.wiestii Steud., A.hirtula Lag., A.atlantica Baum, A.strigosa Schreb. (As genome); tetraploids - A.barbata Pott., A.vaviloviana Mordv., A.abyssinica Hoch. (AB genomes); A.magna Mur.et Terr., A.murphyi Ladiz. (AC genomes); hexaploids - A.fatua L., A.occidentalis Dur., A.sterilis L., A.ludoviciana Dur., A.byzantina Koch, A.sativa L. (ACD genomes).
DNA were extracted from primary leaves of 3-5-weeks-old plant by method (Murray&Tompson, 1980) with some modifications. RAPD analyses was carried out according to protocols (Tsukuba, 1996). For this investigation from 113 previously tested primers were chosen 15 primers only. All of these primers generated 56 scorable amplification products (Table 1.). They were highly informative in the detection of polymorphism between species.
Table 1. Characteristics of primers which were used in RAPD-analyses.
| # | Primer | Structure of primer | Quantity of polymorphic components |
| 1 | 16 | CCGACAGCTT | 3 |
| 2 | 18 | ATGGCCTTTA | 2 |
| 3 | 137 | CAGGCCCTTC | 5 |
| 4 | 140 | GGGTAACGCC | 2 |
| 5 | 151 | GTTGCGATCC | 5 |
| 6 | 196 | GGCTCGTACC | 3 |
| 7 | 207 | GCCTCCTACC | 5 |
| 8 | 208 | GGGGTCGATT | 2 |
| 9 | 212 | GTTAGGTCGT | 5 |
| 10 | 215 | CTGTGCTGTG | 5 |
| 11 | 216 | GGTGATGTCC | 4 |
| 12 | 217 | GGGTTGCCGT | 2 |
| 13 | 223 | CGTTGGATGC | 3 |
| 14 | 262 | CCGATCTAGA | 6 |
| 15 | 271 | CGCATAGGTT | 4 |
| Total quality of components | 56 | ||
Discussion
The most high level of polymorphism was noticed in diploid species (50 components), more less have tetraploids and hexaploids –32 and 26 components respectively. Cluster analysis was carried out on the matrix of DICE coefficient with UPGMA-method. Accessions of different species were discriminated obviously by different ploidy level and genome constitution.
Diploid species consisted of two clusters united variants of A genome (A.longiglumis, A.canariensis, A.wiestii, A.hirtula, A.atlantica, A.strigosa) and C genome (A.clauda, A.pilosa, A.ventricosa) species respectively. Results were in general agreement with morphology based phylogenetic analyses, confirming the large differentiation among A and C genomes in evolution of diploid species. Aside from variants of A genome subdivided into two groups: genomes Ac, Al and As. A certain divergence was observed between two more similarity species A.longiglumis and A.canariensis and other diploid species with A genome (Figure 1). It is agreed with data Nocelli et al., (1999).
{kind=link}
Tetraploid species were divided into two clusters formed by AB genomes (A.barbata, A.vaviloviana, A.abyssinica) and AC genomes (A.magna, A.murphyi). The analyses of tetraploid species relationships confirms the differentiation of the barbata complex from the magna-murphyi group, which, despite some similarities in morphological and biochemical traits, seems to have accumulated deep genetic differences along its evolutionary pathway (Figure 2).
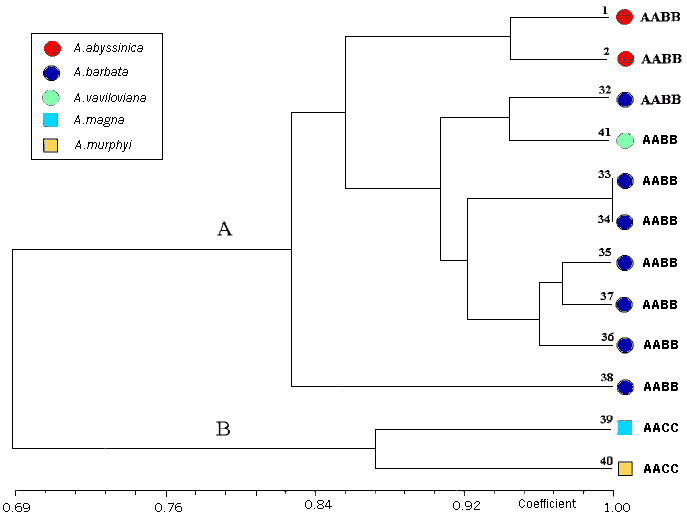{kind=link}
All hexaploid species were grouped into two subgroups, while the diploid species and the tetraploid species showed more divergence. The most visible separate groups have accessions of A.sativa and A.byzantina. In most cases hexaploid species demonstrated the lowest polymorphism by RAPD-markers. It means that all of them have the same structure of genome – ACD. Amplification products specified for A, B, and C genome respectively were revealed (Figure 3).
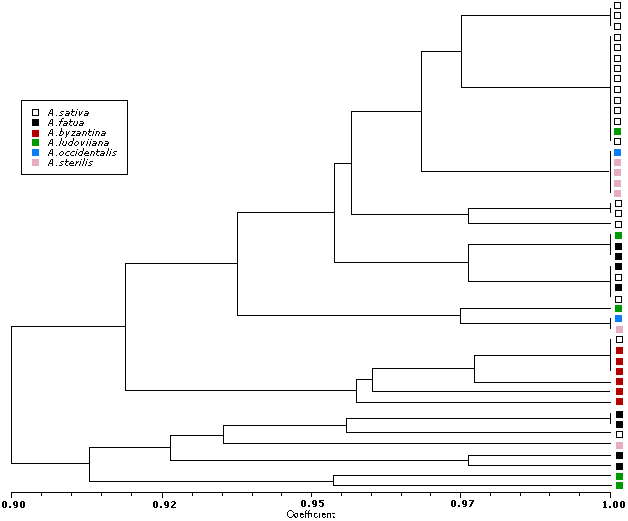{kind=link}
Conclusion
These characteristics could be used as genome markers in further investigations. The given data confirm and add taxonomical system of genus Avena L. (Loskutov, 1999), which are based on morpholigical, karyological and hybridological analyses.
References
- Perchuk I.N., Okuno K., Loskutov I.G., Ebana K. 1999. Interspecies variability of oats by RAPD markers. Proc. XVI International Botanical Congress, USA, p.615.
- Loskutov I.G. 1999. On the taxonomy of genus Avena L. Proc. XVI International Botanical Congress, USA, p.422.
- Nocelli E., Giovannini T., Bioni M., Alicchio R. 1999. RFLP- and RAPD-based genetic relationships of seven diploid species of Avena with the A genome. Genome, v.42, p.950-959.
| Evaluation of interspecific diversity in Avena genus by RAPD analysis | |